Soziale Systeme 1 (1995), H.2,
S. 179-202
Ecosocieties: Societal Aspects of Biological Self-Production
Milan Zeleny
Zusammenfassung: Es wurde mit Erfolg behauptet, daß alle biologischen
(lebenden) Systeme autopoietische Systeme sind. Mit geringerem Erfolg
wurde die umgekehrte These vertreten, daß alle autopoietischen Systeme
biologische (lebende) Systeme seien. Kürzliche Fortschritte auf
den Gebieten des "Artificial Life", der synthetischen
Biologie und der osmotischen Wachstumsprozesse haben aber gezeigt,
daß zumindest einige autopoietische Systeme nichtbiologisch sind:
d.h. autonom und selbstproduzierend in einem physikalischen, anorganischen
Milieu. Das Phänomen Leben ist einer spezifischen Organisation der
Materie zuzurechnen, nicht einer besonderen (e.g. organischen) Materie.
Weiterhin existiert eine ausführliche Diskussion darüber, ob soziale
Systeme (diejenigen, die spontan geordnet und nicht vom Menschen
entworfen sind) autopoietische Systeme sind.
Dieser Essay behauptet, daß die Frage der Autopoiesis spontaner
sozialer Systeme irrelevant ist: Nicht nur, daß spontane soziale
Systeme autopoietisch sein müssen, sondern die Umkehrung dieser
Relation gilt noch bestimmter: Alle autopoietischen und also alle
biologischen (lebenden) Systeme müssen soziale Systeme sein.
Diese These impliziert nicht, daß alle sozialen Systeme autopoietisch
sind; es gibt viele vom Menschen entworfene und gemachte heteropoietische
"Wunder" des sozialen "engineering", die weder
autonom noch selbstproduzierend oder selbsterhaltend sind. Aber
alle autopoietischen Systeme, die organischen und die anorganischen,
müssen notwendigerweise soziale sein (Gesellschaften, Populationen,
Kommunitäten).
Es ist nicht wichtig anzumerken, daß einige (d.h. spontane) Sozialsysteme
autopoietisch sind; entscheidend ist, daß alle lebenden Systeme
(und alle ihre lebenden Subsysteme) Gesellschaften sind und als
solche untersucht werden müssen.
Introduction
One could say that the expulsion of the biological and natural
ecological determinants of human existence has been one of striking
features of sociological studies since the 1930s.
Edward Shils, 1985
E. Shils, in his The Calling of Sociology (1985),
as well as in several later works, insists that the capacity to
discern connections between activities or institutional arrangements
which appear to be unconnected derives from the postulate of the
systemic character of society. Society, according to this postulate,
is a whole of interdependent parts, each of which is the "environment"
for all the others.
Shils's is a properly (in the sense of Smuts) holistic view, recognizing
a whole as a unity of parts that affects the interactions of those
parts. The temporal and territorial parochiality of the traditional
sociological research, although rich in concrete and descriptive
detail, is often short on fulfilling the programmatically announced
intentions of universal validity. Yet, Shils (1985)
insists:
Although sociologists have made little progress in the delineation
of whole societies, either in theory or in particular investigations,
the idea of a "society" as a whole or as a system whose
parts are interconnected in many ways, is a fundamental postulate
of sociology [Italics M. Z.].
Modern sociology is increasingly being challenged by the urgent
issues related to the sustainability or self-sustainability of social
networks (ecosystems, ecosocieties). These challenges can be met
only if the "fundamental postulate of sociology" becomes
firmly embedded within sociological research methodology. Developing
a sufficiently general notion of a "social system," with
sufficiently generalizable organizational and structural properties,
is a necessary step towards producing sound knowledge which can
be of practical utility. The discovery of the fundamental laws of
social life remains an unfulfilled promise of sociology.
According to Shils: "There is at present no systematically
articulated general theory in sociology which would find general
acceptance among sociologists." Consequently, the rich results
of sociological investigations are not clearly and precisely cumulative.
The purpose of this essay is twofold: (1) To propose that many
important social systems are self-producing (autopoietic), rather
than purposefully "engineered" or constructed (heteropoietic),
and, more radically, (2) To propose that all self-producing (autopoietic)
systems, including biological "living" systems, must also
be social systems.
The first proposition attempts to unveil the nature of self-organization
and thus self-sustainability of spontaneous social orders and thus
provide a foundation for non-ideological, non-political and sustainable
social action.
The second proposition suggests and points to potentially significant
intersections of biology, economics and sociology. The resulting
interdisciplinary "crosspollination" of these disciplines
would go a long way towards effectively addressing the issues of
environmental self-sustainability.
Since the times of Parsons and Luhmann, "actions" and
"communications" have been considered the components of
social (sociological) systems. Autopoietic theory attempts to reinstate
the individuals to their proper place as the proper components,
with all their inaction and miscommunication abundant.
Individuals and Networks
Cells move, die, divide, release inductive signals or morphogens,
link to form new sheets, and repeat variants of the process. Genes
control the whole business indirectly by governing which morphoregulatory
or homeotic product will be expressed.
Gerald E. Edelman, Bright Air, Brilliant Fire, 1992
Since Huxley's The Individual in the Animal Kingdom, the
idea of individuality has continued to present fundamental difficulties
in biology. Is a colony of "white ants" an individual?
Huxley proposed that: "Wherever a recurring cycle exists (and
that is in every form of life) there must be a kind of individuality
consisting of diverse but mutually helpful parts succeeding each
other in time, as opposed to the kind of individuality whose parts
are all coexistent." Huxley, in 1912,
distinguished between individuality in time (species-individuality)
and our ordinary, simultaneous or spatial individuality.
It is the first kind of individuality, the individuality of a cyclically
recurring network of "ordinary" individuals, that is crucial
for an enhanced understanding of self-sustaining social systems
or networks.
Every organism, even if spatially and temporarily isolated, can
emerge, survive, and reproduce only as part of a larger societal
network of organisms. Similarly, each cell, organelle, or neuron
can exist only as part of a group or society of cells, organelles,
or neurons. Each component of an autopoietic (Varela/Maturana/Uribe
1974; Zeleny 1981) system
can emerge, persist, and reproduce only within the complex of relationships
that constitute the network of interconnected components and componentproducing
processes.(1)
Before any organism can reproduce, it must first be produced (or
self-produced), and it must survive. Autopoiesis (self-production)
therefore precedes, and in fact creates, the conditions for all
subsequent reproductions.
Survival (economic and ecological) activities of separate organisms
directly form and reform their metasocieties of interactive populations
which are further concatenated into regional networks and full ecosystems.
Reproductive organismic activities can take place only within such
preformed networks and thus assure their own (i.e., networks') reinforcement
and self-production. In fact, autopoietic systems can, and many
do, adapt and evolve without their own reproduction; only their
components may reproduce.(2)
Eldredge (1996) concludes
that a gene-centered view of such systems is unnecessary, and that
social networks are demonstrably biotic systems. The entire human
society can be viewed as such an autopoietic superorganism (Stock/Campbell
1996), embedded in an autopoietic Gaia - as is often propounded
by L. Margulis (Mann 1991).
Margulis has also targeted Neodarwinism(3)
and its inability to answer important questions or explain fundamental
phenomena - for example, there is not a single case of a new species
created by building up of chance mutations. She has embraced the
so called "autopoietic Gaia".
The socalled "Gaia hypothesis" is not new in the history
of science; A. A. Bogdanov formulated it quite clearly (Zeleny
1988a):
The entire realm of life on earth can be considered as a single
system of divergence, based on the rotation of carbon dioxide.
This rotation forms a basis for complementary correlations between
life as a whole - the "biosphere" - and the gaseous
cover of the Earth - the "atmosphere." The stability
of atmospheric content is sustained in the biosphere, which draws
from the atmosphere the material for assimilation.
Bogdanov, the father of tectology (the precursor of modern autopoiesis),
has thus conceptually coupled biosphere, atmosphere, hydrosphere,
and lithosphere into a single holistic(4)
system of mutually co-evolving influences.
Organisms cannot be separated (except through artificial cleavage)
from their economic, ecological, or social environments which they
themselves co-produce and mutually provide to each other. Only a
temporarily disembodied human mind can imagine removing itself,
temporarily, from its social surroundings - from its life base.
Self-Sustainability
The significance of natural ecological and biological qualities
has been denied by sociology in favor of a conception of the environment
as preponderantly, if not exclusively "social". Yet, any
successful addressing of the challenging issues of sustainability
of social systems must address much longer and remoter (past and
future) historical periods than has been customary.
Even though we often talk about sustainable systems, it is the
self-sustainability of systems which is of greater interest. The
question is not: How can we sustain a given system?, but: How can
a system sustain itself in a given milieu?
Another dimension of sustainability must be its organizational
mode. It is important to realize that sustainability (and self-sustainability)
is directly related to system organization and its self-production
(autopoiesis). How are systems organized is much more important
than how its individual agents think or what values they uphold.
Self-sustainable systems are autopoietic and must therefore be
organized for autopoiesis. Sustainable systems are heteropoietic,
i.e., their sustainability does not come from within (from its own
organization) but from without, from planned, system-sustaining
activities of external agents. Non-sustainable systems are allopoietic,
i.e., they are organized to produce things other than themselves.
Allopoietic systems necessarily deplete their environment.
Heteropoietic systems can be sustainable as long as external agents
sustain their system-sustaining efforts. Only autopoietic systems
replenish their own environment and thus can become self-sustaining.
Self-sustainable systems must maintain their ability to coordinate
their own actions. Purposeful coordination of action - or knowledge
- has to be continually produced and maintained: self-sustaining
systems must be knowledge-producing, not just labor or capital consuming
entities. An autopoietic view of knowledge is also presented through
the corporate epistemology of Georg von Krogh et al. (1994).
In summary, the presented view of sustainability can be characterized
as follows: both sustainability and self-sustainability are time
and context dependent system properties emerging from system organization.
System organization must be continually produced or renewed via
operating a common, shared resource system, optimally managed through
competition and collaboration of agents. Continued functioning of
the organization thus requires continued coordination of action,
i.e., continued production of knowledge.
Most systems can be sustained over long periods of time through
an external supporting agent disbursing ideas, effort, money or
resources. Once this external agent withdraws its support, system's
sustainability can be directly challenged. Externally sustainable
systems do not have to be internally self-sustainable.
Any relationship (External agent -> Sustainable system) can
be transformed into a self-sustainable metasystem (External agent
<-> System). While an external agent can in principle make
any system sustainable, only a meta agentsystem can become self-sustainable:
through making the external agent an internal part of the system.(5)
Autopoiesis
If Nature possesses a universal psyche, it is one far above the
common and most impelling feelings of the human psyche. She certainly
has never wept in sympathy, nor stretched a hand protectively
over even the most beautiful or innocent of her creatures.
Eugène Marais, The Soul of the White Ant, 1970
Among the physical, biological, and social systems, the most complex
and the most interesting are those which are autopoietic, i.e.,
autonomous and self-producing. The definition of these systems has
been introduced by Varela, Maturana, and Uribe (1974). Also Haken
(1986), defining synergetics,
refers to systems composed of many individual parts which, by their
cooperation, form organizations and structures - i.e., he refers
to social systems.
An autopoietic system has been defined as a system that is generated
through a closed organization of production processes such that
the same organization of processes is regenerated through the interactions
of its own products (components), and a boundary emerges as a result
of the same constitutive processes.
Varela et al. (1974)
have conceived autopoietic organization as an autonomous unity or
meta-network of productions of components, which participate recursively
in the same network of productions of components, which produced
these components, and which realize such a network of productions
as a unity in the space in which the components exist.
Such organization of components and component-producing
processes remains relatively invariant through the interaction and
turnover of components. The invariance follows from the definition:
if the organization (the relations between system processes) changes
substantially, there would be a change in the system's categorization
in its identity class. What does change is the system's structure
(its particular manifestation in the given environment) and its
parts. The nature of the components and their spatiotemporal relations
are secondary to their organization and thus refer only to the structure
of the system.
System's boundary is a structural manifestation of the system's
underlying organization. The boundary is a structural realization
of the system in a particular environment of components. In physical
environments this could take the form of a topological boundary.
Both organization and structure are mutually interdependent.
The concepts of the autopoietic nature of a system were developed
by Varela et al. (1974)
based on a living (biological) system as a model of self-production.
Yet self-production has the potential to mean and be interpreted
through many different ways by a variety of observers. "Autopoiesis"
has been coined (not translated from Greek) as a label for a clearly-defined
interpretation of "self-production." This phenomenon of
self-production can be observed in all living systems. A cell, a
system that renews its macromolecular components thousands of times
during its lifetime, maintains its identity, cohesiveness, relative
autonomy, and distinctiveness despite such intense turnover of matter.
This persisting unity and its holism is called "autopoiesis."
Zeleny (1981) presents
an overview of autopoiesis as a theory for the living organization.
Varela et al. (1974)
have developed a six-point key that provides the criteria for determining
whether or not a system is autopoietically organized.
These criteria, as they are applied to biological (living) systems,
can also be applied to other systems that are currently not considered
"living". This is a simple exercise with very important
implications. We have found (Zeleny/Hufford
1991; 1992) that
not only are spontaneous social systems autopoietic, but also that
the relationship is much stronger. Although all living systems are
autopoietic, not all autopoietic systems are living. For example,
inorganic osmotic growths (Zeleny/Klir/Hufford
1989) are often temporarily autopoietic.
All autopoietic systems must be social systems. In other
words, all autopoietic, and therefore all biological (living) systems,
are social systems. Also, the topological boundary, that has been
necessary to describe an autopoietic system within a favorable environment
of physical components (such as those within and around a cell),
may not necessarily take a physical form in other types of systems,
e. g., in social systems.
In social systems, dynamic networks of productions are being continually
renewed without changing their organization, while their components
are being replaced; perishing or exiting individuals are substituted
by the birth or entry of new members. Individual experiences are
also renewed; ideas, concepts and their labels evolve, and these,
in turn, serve as the most important organizing factor in human
societies. The organizing core for the implementation of ideas must
be the emergent society as an autopoietic entity.
Autopoietic systems can persist in their autopoiesis for many decades
(humans, trees), for many days (cells) or for mere flashes of hours,
minutes, seconds, or milliseconds (osmotic growths). The "lifespan"
of autopoiesis in no way enters (or should enter) into its definition.
Also, autopoiesis is bound to exhibit gradation; it does not jump
into being in a magic instant - it becomes. It gradually degrades
itself; the processes of autopoiesis weaken and dim, more or less
rapidly (Zeleny 1978).
There is a great modeling, methodological and explanatory potential,
certainly on the rise in modern sciences, in treating autopoietic
systems as social systems.
Social Systems
"Have you ever seen, in some wood, on a sunny quiet day,
a cloud of flying midges - thousands of them - hovering, apparently
motionless, in a sunbeam? ... Yes? ... Well, did you ever see
the whole flight - each mite apparently preserving its distance
from all others - suddenly move, say three feet, to one side or
the other? Well, what made them do that? A breeze? I said a quiet
day. But try to recall - did you ever see them move directly back
in the same unison? Well, what made them do that? Great human
mass movements are slower of inception but much more effective."
Bernard M. Baruch, Foreword to Mackay's Extraordinary Popular
Delusions, 1849
It is time to define social systems and to elucidate the meaning
of "social" for the purposes of this paper. Such definition
should be as general as possible, encompassing a rich variety of
contexts, yet being fully amendable, adjustable and applicable to
all such contexts.
Social systems are renewable, self-producing networks characterized
by internal (rather than external) coordination of individual action
achieved through communication among temporary agents. The key
words are coordination, communication, and limited lifespan of the
agents or their groupings.
It should be self-evident that the general notions of coordination,
communication and individual lifespan will acquire different meanings
in different contextual embeddings.
Coordinated behavior includes both cooperation and competition
(and all forms of conflict), in all their shades and degrees. Actions
of predation, altruism, and self-interest are simple examples of
different and interdependent modes of coordination. Communication
could be physically, chemically, visually, linguistically, or symbolically
induced deformation (or in-formation) of the environment and consequently
of individual action taking place in that same environment.
So I, as an individual, can coordinate my own actions in the environment
only if I coordinate it with the actions of other participants in
the network. In order to achieve this, I have to in-form (change)
the environment so that the actions of others is suitably modified;
I have to communicate. As all other individuals are attempting to
do the same, a social network of coordination emerges, and, if successful,
it is being "selected" and persists. Such a network improves
my ability to coordinate my own actions within the environment effectively.
Cooperation, competition, altruism, and self-interest are thus inseparable.
Social systems cannot and should not be limited to human systems,
especially when addressing the issues of ecological self-sustainability.
Human systems simply in-form a special meaning on the universal
acts of coordination, communication, and birthdeath processes in
general social systems.
A group of fish thrown together by a tide wave is a passive aggregation,
not a social system. A swarm of moths lured to a porch light is
an active aggregation, but not a social system. A flagpattern of
athletes "constructed" through bullhorn-shouted commands
from a coordination center is a purposeful heteropoietic aggregation,
not a social system.
All of these can transform into social systems as soon as internal
communication patterns become established; they should then temporarily
persist (become autonomous), even after removing the external impetus.
Merely externally induced interaction of components does not suffice:
billiard balls interact and so do wind-blown grains of sand - nobody
would call them social systems.
Human waiting queues are often engineered and externally induced
(enforced, not voluntary) interactions. To a large degree however,
they do exhibit, at least temporarily, the voluntary self-organization
characterized by its own specific behaviors, rules of conduct, choice
of distance and modes of communication.
Similarly, schools of fish, swarms of bees, flocks of birds, packs
of animals, and even the spontaneous wavepatterns of Olympic-games
spectators are, however, no matter how ephemerally shortlived, undoubtedly
social systems.
Any social system, in order to adapt and persist in its environment,
must be capable of reshaping itself, controlling its growth, and
checking the proliferation of individuals. In other words, the long-term
persistence of a social system is critically dependent on harmoniously
balanced birth and death processes. There can be no collective life
without individual death.
A proliferation of individuals without balancing death processes
and without deathinducing communication is "cancer" -
a shortlived, environmentally destructive outburst of life-like
processes, but not the life itself. A dominant death process, without
a sufficient birth-process complement, takes any social system towards
its extinction.
Life of a social system, and thus life itself, is based on a dynamic
and autopoietic harmony between birth and death processes. Life
is necessarily a social phenomenon: the life of an individual cannot
take place outside a social network, and the individual life itself
must be socially embodied at the level of its components.
This view is quite different from the deterministic and essentially
non-biological dogma that (somehow) the growth of an organ is genetically
(symbolically) programmed into the cells which are then guided (read-only
memory) by this "geneprogram" through an exquisitely precise
and predetermined series of events.
No communication and no death implies no life.
Detecting Autopoiesis in Systems
To illustrate the diversity of autopoiesis in its application to
systems analysis, Zeleny and Hufford (1991;
1992) have analyzed
three systems: a biological (living) system, a chemical system,
and a spontaneous social system. Here we summarize their main conclusions.
1. The Eukaryotic Cell
The generalized non-plant eukaryotic cell may be described as having
a plasma membrane which surrounds the cytoplasm and cytoplasmic
components of the cell. The cytoplasm contains the nucleus, mitochondria,
golgi apparatus, endoplasmic reticulum, various vesicles, lysosomes,
vacuoles, cytoplasmic filaments and microtubules, centrioles, and
other components of the cell.
After applying the Varela-Maturana-Uribe six-point key to the generalized
eukaryotic cell, it can be concluded that the cell is an autopoietic
unity in the space in which its components exist.
L. Margulis (Mann 1991) is one
of the few biologists who viewed eukaryotic cells as autopoietic
populations of components. "We are walking communities,"
she insisted.
2. Osmotic Growth
Stephane Leduc (1911) described
an "osmotic growth," a membrane of precipitated inorganic
salt, as having many processes, functions, and characteristic forms
that appear to be analogous to those found in living systems. The
osmotic experiments performed by Leduc have been also reproduced
by Klir, Hufford, and Zeleny (1988).
Unlike typical experiments in simple precipitation, where two solutions
are mixed and a cloudy solution of an insoluble salt results, osmotic
growths precipitate and grow over a period of minutes to days and
go from a thin transparent membraneous state to an opaque state.
An actual photographic sequence has been provided by Zeleny, Klir,
and Hufford (1989).
After applying the six-point test, based on the evaluation of osmotic
growths (specifically the calcium chloride/tribasic sodium phosphate
system), it can be concluded that an osmotic growth is an autopoietic
unity in the space in which its components exist.
At the macroscopic level, the osmotic precipitation membrane exhibits
fluidity, elasticity, and resealability identical to the properties
of the plasma membrane. As the internal osmotic pressure increases,
an expansion occurs (not a rupture) allowing components from the
internal and external spaces to flow through the membrane and "couple"
within the membrane. The osmotic growth phenomenon occurs because
the operational integrity of the precipitation membrane is maintained.
Osmotic growths are, temporarily and often ephemerally, autopoietic.
This implies that if we hold the current autopoietic theory to be
correct and intact, then we must reassess our definition (redefine
our criteria) of what it means to be "living". If we do
not give up our current definitions of "living", then
we must conclude that there is a fundamental problem within the
existing theory of autopoiesis which needs to be addressed.
3. Kinship System - A Spontaneous Social System
As our third system, the kinship system is an example of a spontaneous
social order that has a substantial impact and great significance
in the life of social, economic, and political networks. A kinship
system constitutes, prototypically, an autopoietic system that is
produced and maintained through organizational rules (which are
potentially codified) of a given society. No matter what the particular
mix of its components (men, women, and children), the kinship system
organizes its social domain and coordinates its social action in
a spontaneous, self-perpetuating fashion. It must continually adapt
to the external challenges and interferences of the society,(6)
social engineers and reformers.
Social networks, embodying kinship systems, are not static and
unchanging structures, but highly dynamic ones. Cochran et. al.,
in their study of kinship systems (1990),
established that the distribution of different types and roles of
network participants (kin, friends, neighbors, formal ties) remains
relatively stable, even though the names and faces of network members
keep changing. In the language of autopoiesis: It is their organization
that remains stable, while their structures and components continually
change.
Social networks can therefore change in their structure or in the
nature of their component relationships (organization). One can
therefore study shifts in the network's structure, turnover among
its members, and changes in the character of continuing network
ties. For example (Cochran et al.
1990), in spite of frequent moving and changes of neighborhoods,
American white children maintain the largest stable social networks
(8 adults, 8 peers) while relatively immobile Swedish children maintained
the smallest (4 adults, 4 peers).
Viewing families and kinship networks properly as autopoietic systems
could lead to new and important understanding of the effects of
residential mobility, divorce rates, death and disease disruptions,
loss of employment, or state intervention on the structure, organization
and durability of social bonds in important social and support networks
- primary, functional, peripheral and formal.
Through social autopoiesis, one also can learn more about which
social environments produce desirable social supports in transaction
with parents. What is the role of friends and relatives? What is
the role of parental self-confidence, and how can it be enhanced?
What is the role of a parent's level of formal education? How do
intervention programs interact with the spontaneous self-organizational
nature of social autopoiesis? The research agenda of self-producing
social systems is remarkable in its challenge and significance.
It was F. A. Hayek who integrated the concepts of self-production
directly into the domain of social systems (1988).
Hayek stated that:
Although the overall order of actions arises in appropriate circumstances
as the joint product of the actions of many individuals who are
governed by certain rules, the production of the overall order
is of course not the conscious aim of individual action since
the individual will not have any knowledge of the overall order,
so that it will not be an awareness of what is needed to preserve
or restore the overall order in a particular moment but an abstract
rule which will guide the actions of the individual.
Consequently, the individuals in a society spontaneously assume
the sort of conduct and evolve the rules which assures their continued
existence within the whole. Of course, this conduct and rules must
also be compatible with the preservation of the whole. Neither the
society nor the individuals could exist if they did not behave in
this manner. The overall order, preservation of the society, is
not the "purpose" or the "plan" of the individuals.
The individual actions are motivated by their own goals and purposes.
Amoeba: Biotic Social Systems
What do human beings, ants, and slime have in common? Despite
their differences in structure, physiology and ecology, all three
consist of individuals whose behavior is sufficiently coordinated
for the group to be called a society.
Howard Topoff, 1981
Is this "coordination" and the resulting society due
to executing a preconceived plan of a social engineer, central planner,
or a great designer (like in heteropoietic systems), or is it due
to the distributed and unintended self-coordination of goalseeking
and autonomously behaving individuals (like in autopoietic systems)?
Cellular Slime Mold (Garfinkel
1987) is another good example of an autopoietic social system.
The slime molds (Gymnomycota) are an example of a fungus-like protist.
They are decidedly fungus-like at some stages and animal-like at
others. Their life cycle includes an ameoboid stage and a sedentary
stage in which a fruiting-body develops and produces spores.
In Dictyostelium discoideum (Garfinkel
1987), the vegetative cell is ameoboid. Ameobas are individual
cells moving around in search for bacteria to feed on. They will
grow and divide indefinitely. Often they digest so much and produce
new amoebas so rapidly that their food supply has no chance to replenish
itself. When the food supply has been exhausted, they move rapidly
to a central point, collecting themselves into a well-differentiated
spontaneous aggregation (center cells, boundary cells, etc.) - a
pseudoplasmodium. The aggregation is triggered by the production
of cyclic adenosine monophosphate (AMP) which attracts other amoebas
in a chemotactic fashion.
The group then assumes the shape of a "slug" with a head,
tail, and an apparent "purpose": searching collectively
for a new, potential source of food. Around the outside is secreted
a mucoid sheath (aggregate boundary). It migrates as a unit across
the substratum as a result of the collective action of the amoebas.
The changing of the roles of individual amoebas is prevalent; the
original leaders who formed the center of attraction are dispersed
throughout the "slug", and new leaders emerge, forming
the "goalseeking" head.
The head of the homehunting "slug" are simply the fastest-moving
amoebas. The "slug" is just a spontaneous temporary metaorganism,
preserving each amoeba as a separate individual. The slug is positively
phototactic (migrates toward light), and it usually migrates for
a period of hours. Its behavioral responses are essential "to
ensure" that the spores will be borne in the air and so can
be effectively dispersed.
Fruiting body formation begins when the slug ceases to migrate
and becomes vertically oriented. The "leading" amoebas
change quickly from the first to the last. The head of the slug
forms the base of a stalk which follower-amoebas continue to build
(they secrete cellulose to provide rigidity) up into a mushroom-like
metaorganism. At its top, hundreds of thousands of amoebas differentiate
into spores that are embedded in slime and, after the mushroom "head"
matures, it bursts. It disperses the spores to new and potentially
nourishing environments. When they fall to earth, they change once
again into the individual amoebas which reproduce by cell division.
This ecological cycle is then repeated.
Amoeba: Human Social Systems
To the naive mind that can conceive of order only as the product
of deliberate action, it may seem absurd that in complex conditions
order, and adaptation to the unknown, can be achieved more effectively
by decentralising decisions, and that division of authority will
actually extend the possibility of overall order.
F. A. Hayek, The Fatal Conceit, 1988
After the undisputed failures and fatal conceit of large-scale
social engineering and experimentation of the past (Hayek 1975;
1988), the phenomena of spontaneity and emergence in social systems
are becoming important again. The survival and robustness of social
institutions, such as market, family, culture, money, language,
economy, city, and myriads of other voluntary orders, are being
noticed.
The biological amoeba metaphor has recently found its organizational
embodiment in the most successful "amoeba system" at Kyocera
Corporation (Hamada/Monden
1989).(7)
The Kyocera "amoebas" are independent, profit sharing
and self-responsible units of three to fifty employees. Each amoeba
carries out its own statistical control, profit calculus, cost accounting
and personnel management. They compete, subcontract, and cooperate
among themselves on the basis of intracompany market of transfer
prices.
Depending on the demand and amount of work, the amoebas can divide
into smaller units, move from one section of the factory to another,
or integrate with other amoebas or departments. All amoebas are
continually on the lookout for a better buyer for their intermediate
products. Many amoebas even produce the same or similar products.
They are authorized to trade their intermediate products with the
outside companies; if the internal vendor is unreasonable, the buyer
amoeba will search for a satisfactory supplier outside the company.
A most remarkable feature towards system autonomy is the member
trading. Heads of amoebas lend and borrow members and so eliminate
losses caused by surplus labor. So, Kyocera's amoebas multiply,
disband, and form new units in the spirit of autopoiesis (self-production)
of the enterprise. Amoeba division and breakup are frequent occurrences
and are based on the criteria of output and a worker's added value
per hour.
This concept of ultimate flexibility is best summed up by Kyocera's
President Inamori: "Development is the continued repetition
of construction and destruction" (Hamada/Monden
1989), as if coming directly from the systems theories of autopoietic
self-organization.
Another example would be Australian TCG (Technical Computer Graphics),
a self-producing network in a business-firm environment. There are
no coordinating divisions, "leading firms", or management
superstructures guiding TCG's 24 companies; the coherence, growth
and maintenance of the network is produced, according to J. Mathews
(1992), by a set of network-producing
rules.
Boundaries of Social Systems
In kinship systems, their boundaries are usually well defined.
The distinction between family and non-family members is rarely
ambiguous or subject to fuzzy interpretation. A definite family
boundary can be established, although it is not necessarily topological.
In the context of the family, the concept of boundary might be defined
as the members included in a set. Family members are usually distinguished
from their environment (from the "society") more sharply
than any engineered or designed physical "membrane" can
assure.
All social systems, and thus all living systems, create, maintain,
and degrade their own boundaries. These boundaries do not separate(8)
but intimately connect the system with its environment. They
do not have to be just physical or topological, but are primarily
functional, behavioral, and communicational. They are not "perimeters"
but functional constitutive components of a given system.(9)
Boundaries do not exist for a human observer to see or identify
the system, but for the system and its components to interact and
communicate with its environment. Boundaries range from phospholipid
bilayers, globular proteins, osmotic precipitates, and electric
potentials, through cell layers, tissues, skins, metabolic barriers,
and peripheral neural synapses, to laterally or upwardly dispersed
boundaries of territorial markers, lines of scrimmage, social castes,
secret initiation rites, and possessions of information, power,
or money.
A company can have a number of geographically separate offices
or be a virtual company, entirely "in the air" of electronic
communication. The U.S.A. includes Alaska and Hawaii. A doctor does
not leave the social system of a hospital while "on call"
or connected with a beeper. Many additional examples and details
of non-topological social boundaries are discussed by Miller and
Miller (1992).
Although social systems are necessarily physical because their
components realize their dynamic network of productions in the physical
domain (their components are cells, termites, lions, adult humans,
etc.), many computer simulations (Zeleny
1978) of autopoietic systems show that topological boundaries
arise only if very minute rates of production processes are very
finely adjusted and harmonized. In other words, the underlying organization
of processes has to be "tuned up". If not, a human observer
might not be able to "see" or recognize any "topological"
boundary. Yet the organization remains functional and invariant;
autopoiesis continues; we do not see any boundary, but the system
remains autopoietic.
All Autopoietic Systems are Social Systems
Recent advances in the areas of Artificial Life (Langton 1989),
synthetic biology, and osmotic growths (Klir/Hufford/Zeleny
1988; Leduc 1911; Zeleny/Klir/Hufford
1989) have established that at least some autopoietic systems
could be nonbiological, i.e., self-producing in inorganic milieus.
Autopoiesis can take place only where there are separate and autonomously
individual components interacting and communicating in a specific
environment according to specific behavioral (including birth and
death) rules of interaction.
Approaches which sacrifice this essential individuality of components,
like the statistical systems of differential equations used in the
traditional systems sciences, cannot model autopoiesis. They are
definitionally incapable of treating autopoietic systems as social
systems. Components and participants in autopoiesis must follow
rules, interact, and communicate - they must form a community of
components, a society: a social system.
F.A. Hayek (1988) pointed
out that social engineers assume that since people have been able
to generate some systems of rules coordinating their efforts, they
must also be able to design an even better and "improved"
system. The traditional norms or reason guiding the imposition and
subsequent restructuring of socialism embody a naive and uncritical
theory of rationality, an obsolete and unscientific methodology
which Hayek calls "constructivist rationalism" and which
E. L. Khalil (1990) traced to
Karl Marx's concept of social labor.
Although the family (and other spontaneous social orders (Zeleny
1985; 1991)) can easily
produce and generate systems other than itself, its primary capability
is that of producing (and reproducing) itself.
The removal of external pressures, support and props is one of
the safest tests of viability (i.e., autopoiesis) in social systems.
If the coercive boundaries (physical or otherwise) dissolve, and
the social system ceases to exist, it was not autopoietic; if it
reasserts its social boundary and voluntarily increases the level
of cohesiveness, then it is autopoietic and self-sustaining.
It is only in the sense of such centrally-imposed "command"
systems that we present our conjecture: All autopoietic (biological)
systems are social systems.
Social organization can be defined as a network of interactions,
reactions, and processes involving at least:
1) Production (poiesis): the rules and regulations guiding the
entry of new living components (such as emergence, birth, membership,
acceptance).
2) Bonding (linkage): the rules guiding associations, functions,
and positions of individuals during their tenure within the organization.
3) Degradation (disintegration): the rules and processes associated
with the termination of membership (death, separation, expulsion).
In Figure 1 we graphically represent the above three poietic processes
and connect them into a cycle of self-production. Observe that all
such circularly concatenated processes represent productions of
components necessary for other processes, not only the one designated
as "production..". To emphasize this crucial point we
speak of poiesis instead of production and autopoiesis instead of
self-production. Although in reality hundreds of processes could
be so interconnected, the above three-process model represents the
minimum conditions necessary for autopoiesis to emerge.
From the vantage point of Figure 1, all biological (autopoietic)
systems are social systems. They consist of production, linkage,
and disintegration of related components and component-producing
processes. An organism or a cell is, therefore, a social system.
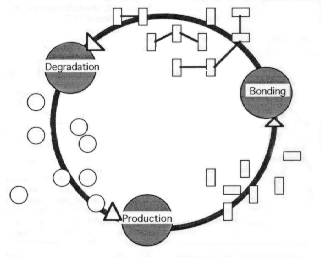
Figure 1: Circular organization of interdependent processes
and their "productions"
Marvin Minsky has titled his recent book The Society of Mind
(1986), attempting to exploit
the social metaphor in studying the mind as a society. According
to Minsky, mind is neither a unified, homogeneous "black box"
or entity, nor a collection of entities, but a heterogeneous system
of networks of processes. Unfortunately, Minsky's view of "society"
is the hierarchy of agents (or experts), based on extreme division
of labor (Zeleny 1988b),
each of them doing "some simple thing that needs no mind or
thought at all."
Minsky writes like a social engineer of command systems, with little
or no awareness of spontaneous social orders (Zeleny
1985): " ... when we [italics M. Z.] join these
agents in societies - in a certain and special way - this leads
to true intelligence."
G. M. Edelman (1988) improves
upon Minsky by stating:
Any satisfactory developmental theory of higher brain function
must remove the need for homunculi and electricians at any level
and at the same time must account for object definition and generalization
from a world whose events and "objects" are not prelabeled
by any a priori scheme or top-down order.
Biological Organisms As Social Systems
The body of a mammal with its many vital organs can be looked
upon as a community with specialized individuals grouped into
organs, the whole community forming the composite animal.
Eugène Marais, The Soul of the White Ant, 1970
Although here we cannot analyze biotic systems in specialist's
detail, let us explore the cellular organism, including the human
organism, as a social system. Living organisms have often been studied
as "black boxes," or as components-free machines, by mechanistic
cybernetics.(10)
Biological organisms are not components-free black boxes but communicating,
birth-death process balancing social systems. Jim Michaelson of
Harvard is one of the few biologists who is prepared to treat biological
systems as social systems, positing the "competition"
of cells, the selection and survival of the most "fit"
during their embryonic development, as being dependent on the cell's
ability to secrete enzymes, rates of proliferation, etc.
1. Communication
Whenever a living cell is unable to communicate with other cells,
it does not die, but rather grows uncontrollably, multiplying into
other noncommunicating cells, forming a malignant tumor which is
unable to survive in its life-sustaining environment because it
destroys it.
All organismic cells are interconnected through tiny channels in
cell membranes or gap junctions. Through these channels, all molecular,
chemical, metabolic, and electric communication among cells takes
place. These communicative junctions are made of proteins (connexins)
that align all cells into one continuous channel-network: a social
system.
Malfunction in intercellular communication channels affects the
intercellular social system and thus could "kill" the
organism itself. If regulatory and inhibitory signals do not get
through, the uncontrolled, deathless growth, and the voracious feeding
on its own environment, would result.
To study cancer processes without studying cellular gap junctions
amounts to a case of professional neglect. Clogged channels block
social-regulatory signals and allow cells to go awry; clear channels
allow the propagation of deadly signals. Gap junctions themselves
are selective self-regulatory; they tend to close and protect against
chaotic signals and to open for and receive regulatory signals.
Even a fetus could not develop if particular groups of cells would
not stop reproducing and growing "just-in-time", or more
precisely, would not start dying.
In order to treat cancers, one has either to re-establish communication
channels and thus self-regulation or block the growth of communication
and support channels (like blood capillaries) in order to stop rampant
proliferation. This is not a trivial mechanistic task; it can be
mastered if we view biological systems as social systems.
2. Social Neighborhoods
As discussed in an AAAS symposium volume (Zeleny
1980), cellular neighborhoods, rather than some inheritable
genetic "programs", are the main determinants of cells'
functions. Sociologically, autopoietic systems are better illustrated
by the American plan of development, where one's status and fate
are determined by one's neighborhood, rather than by the British
plan, where one's status and fate are determined by one's ancestors.(11)
The neural network especially, i.e., autonomous autopoietic system
embedded in a larger complex of organismic networks, requires quickresponse
flexibility and adaptability which cannot wait for a mutations buildup
or rely on requisite but cumbersome "genetic alterations".
Neural networks develop as autopoietic societies; individual cells
wander around, get exposed to differential signalings of different
cellular neighborhoods, and ultimately settle down (or get captured)
within these neighborhoods, becoming functioning neurons of the
visual, hearing, or smell regions of the cerebral cortex.
H. Maturana insists in (Zeleny
1980) that "genes" and viral DNA are only structural
components of autopoiesis. Their distribution and mutation therefore
affect structures and structural characteristics (inheritable shapes
and adhesion properties of proteins), but they do not partake in
organization: they do not organize matter, but are themselves organized
and ordered by autopoiesis.
The greatest error biologists could make at this paradigmatic bifurcation
point is searching for the seat of the master plan behind the body's
gray matter. There is no master plan and there are no black-box
feedback loops within feedback loops. There is only a society in
autopoiesis, organizing matter of different structural attributes
and properties (including viral DNA), thus arriving at different,
sometimes important, structural manifestations. Dr. C. L. Cepko
of Harvard Medical School put it quite bluntly: "The mother
cells do not impart specific information to their daughters about
what to become."
3. Death Process
In addition to communication, social systems are also characterized
by limited lifespans of individual agents-components, i.e., by death.
If molecules would not break down, or cells, organisms, individuals
and entire species would not die, there would be no social systems
and thus no self-sustaining life on Earth.
Death dominates development. The vestigial webbing between human
fetus fingers must be dissolved before birth. About eighty percent
of the nerve cells of the baby's brain must perish within hours
of their creation. Caterpillar's crawling muscles must be sloughed
off in order to have a butterfly; female genitalia must be whittled
away in order to have a male.
Still, an uncontrolled and massive death is non-redeeming: Alzheimer's,
Parkinson's, and Lou Gehrig's degenerative disorders result. Uncontrolled
and massive birth is equally unredeeming: cancerous cellmasses,
killing their own environment (i.e., host organism) result. Individuals
must die in order to maintain their social system.(12)
Death is not a chaotic, haphazard, or disorganized part of social
system autopoiesis; it is a harmonized, choreographed, and often
suicidal dance of the most exquisite complexity. The creation of
autopoiesis is inconceivable without the trimming of apoptosis.
The study of apoptosis is crucial in biology: in fact, no true
biology can exist without it.(13)
Death is not the absence of life, but the crucial building block
of life. Life is never "individual" life, but life of
a social network of balanced and communicating birth-death processes.
A good example is the immune system. Millions of T and B cells
are continually generated, each capable of assaulting foreign proteins,
but unfortunately also the body's own proteins. Up to 98 percent
of them have to undergo immediate apoptosis in order to maintain
the body's autopoiesis in a hostile environment.
Death is a productive process of the social system; it creates
space, it generates production substrate, it brings in the innovation,
and it allows trial-and-error adaptation to the environment. Individual
cells are created in order to die, and thus their social system,
i.e. living organism, can persist.
4. Evolution
The idea that reason, itself created in the course of evolution,
should now be in a position to determine its own future evolution
is inherently contradictory, and can readily be refuted.
F. A. Hayek, The Fatal Conceit, 1988
Social systems persist. They can persist as societies of agents
only if their individual agents are born, communicate, and die in
harmony with themselves and their environment. Because of the turnover
of components, the social networks not only persist and are renewed,
but they also evolve.
The unit of evolution (at any level) must therefore be a network
capable of variety of self-organizing configurations. It is the
entire social network, including neuronal groups (Eldredge
1996), that is being "selected", not its individual
components. Such evolving networks are interwoven and co-evolving
with their environment; they do not only adapt to the environment,
but also adapt the environment to themselves - through mutually
intimate structural coupling.
A bird must undoubtedly adapt to a mountain. However, a society
(network) of birds can make the mountain adapt to them. By overconsuming
particular berries, the new brush growth is controlled, the mountain's
erosion enhanced, and the production of both berries and birds thus
limited until a temporary balance or harmony is restored. Colors
of flowers have coevolved with the trichromatic vision of bees;
shapes of flowers with the structural traits of insects and animals;
modern breeders with the changing tastes and preferences of man.
We quote from R. Lewontin (1982):
The environment is not a structure imposed on living beings from
the outside but is in fact a creation of those beings. The environment
is not an autonomous process, but a reflection of the biology
of the species. Just as there is no organism without an environment,
so there is no environment without an organism.
Varela et. al. (1991),
in their book The Embodied Mind,(14)
conform to the view that living beings and their environments stand
in relation to each other through mutual specification or codetermination:
The world is not a landing pad into which organisms parachute;
nature and nurture stand in relation to each other as product
and process.
This new view of evolution of social networks implies that there
can be no intelligent distinction between inherited and acquired
characteristics. What evolves is neither genetically encoded nor
environmentally acquired, but is ecologically embedded in a social
network. The sociological implications of such realization are profound.
There is also no one fixed or pregiven world (a universe), nor
is its dynamics simply observed or viewed differentially from a
variety of vantage points (a multiverse), but this world itself
is continually re-shaped, and re-created by coevolving social networks
of organisms.
The evolution of paradigms is itself an autopoietic process, and
it is thus inevitable to see how the aged "revolutionaries"
are clinging to the old and suddenly ineffective ideas, how they
themselves have become conservatives, and how they individually
resist the new interpretations of their younger colleagues, often
without realizing that their collective time has passed ...
Closure
When I began my work I felt that I was nearly alone in working
on the evolutionary formation of such highly complex self-maintaining
orders. Meanwhile, researches on this kind of problem - under
various names, such as autopoiesis, cybernetics, homeostasis,
spontaneous order, selforganization, synergetics, systems theory,
and so on - have become so numerous...
F. A. Hayek, The Fatal Conceit, 1988
Living systems, i.e., cells, organisms, groups, and species are
social systems. Their interaction forms the entire terrestrial biosphere
or Gaia, a social system akin to the unified organism of a living
cell, which itself is a social system of its constitutive organelles.
Connecting different species into a coherent, interactive, and
self-organizing system cannot happen without the death and dying
- the fuel of environmental adaptation. The natural death of species
does not signal maladaptability of the species, but harmony, adaptability,
and systemic perseverance of the social network of species. Death
is a cosmological event - the most exquisite assurance of life yet
to be. At one point, individuals of all species receive, by waves
on the shore, sound of the wind, or with radio telescopes, the exquisite,
life sustaining message: "Now, now it would be indecent not
to die."
Harmony and fitness does not imply dominance or competitive advantage
but intimate coupling with the environment through all-embracing
communication. The nature, as a social system, is replete with communication
channels of great variety and subtlety. All life on earth (and most
likely interstellar too) is interconnected through internal and
external harmonies, often unnoticed or ignored by linear sciences.
The connexins of cells, dances of bees, odors of fire ants, allochemicals
of Douglas firs, and the language of humans are only the hints,
only the shy peepholes into the veiled mysteries of life - and the
promises of science yet to come.

|
